Notes RBG Edinb. 45(3): 527-548 (1988)
|
NOTES Royal Botanical Garden Edinburgh
Notes RBG Edinb. 45(3): 527-548 (1988) |
|
ABSTRACT. A study of the intergeneric relationships in the family Pinaceae based on the morphology of a variety of reproductive and vegetative parts indicates the presence of eleven genera arranged in four subfamilies. The subfamily Piceoideae Frankis, subfam. nov. is proposed, and the circumscriptions of subfamilies Laricoideae Melch. & Werd. and Abietoideae Pilger are emended.
The family Pinaceae contains a total of ten commonly accepted genera (Rushforth, 1987; Krüssmann, 1985; Dallimore & Jackson, 1966): Abies, Cathaya, Cedrus, Keteleeria, Larix, Picea, Pinus, Pseudolarix, Pseudotsuga and Tsuga, while Page (1988) accepts a further two genera, Nothotsuga and Hesperopeuce. The inter-relationships of these ten to twelve genera have been discussed in the past on a number of occasions, primarily on the basis of leaf arrangement, the presence or absence of short shoots, and the petiole and pulvinus form, culminating in the subfamily arrangements proposed by Pilger (1926) and Melchior & Werdermann (1954), as adopted by Cheng & Fu (1978), Krüssmann (1985) and others, outlined below (after Krüssmann, 1985):
These genera show however a much wider range of differing and shared characteristics than is indicated by this classification. It proves in particular very unsatisfactory with regard to the morphology of the cone and seed, which appear to have been discussed surprisingly little in view of the traditional importance attached to reproductive morphology in classification systems since Linnaeus's time.
The purpose of this paper is to consider the inter-relationships and generic boundaries in the Pinaceae on a wider basis and with particular reference to the reproductive morphology. Care has been taken to give special emphasis to features which the author considers to be unlikely to have been affected by evolutionary pressures and which therefore represent genuine relationships as opposed to convergent evolution.
In order to carry out this project, fresh and herbarium material of an extensive selection of species of ten of the above twelve genera has been studied, as has herbarium material only of Nothotsuga. It has unfortunately not been possible to study material of Cathaya, to which the author has been limited to published illustrations and descriptions in Chun & Kuang (1958) and Cheng & Fu (1978); conclusions with regard to this fast genus must therefore be regarded as tentative.
Eleven genera are accepted in Pinaceae here. Since the work of Carrière (1867) no significant questions have been raised as to the precise circumscriptions of the genera Cedrus, Keteleeria, Larix, Picea, Pseudolarix and Pseudotsuga; I have found no further evidence to change this view and their circumscriptions are upheld. The remaining five genera include problematical species or are open to alternative interpretations, summarized briefly below. Reasons behind their circumscriptions here are detailed under the discussions of inter-generic relationships.
The genus Pinus is very large, variable and complex, and four segregate genera have been named, Strobus Opiz, Cembra Opiz, Caryopitys Small and Ducampopinus Chevalier. An extensive study into the relationships in Pinus and the possible acceptance of these genera is in progress (Frankis, in prep.); in the meantime the traditional broad circumscription is retained.
One species of Abies, A. bracteata D. Don, differs greatly from all other Abies species; although it has not at any time been given separate generic recognition, there may be good grounds for doing so. The preference of this author is however to retain it in Abies as a distinct sub-genus, as treated by Liu (1971) and others.
Cathaya remains a very poorly known genus. Its single species C. argyrophylla Chun & Kuang (Cheng & Fu, 1978) has been referred to Tsuga by de Laubenfels & Silba (Silba, 1984) though on very tenuous evidence, and also to Pseudotsuga by Greguss (1970; nom. inval. [basionym not fully cited]) on the basis of its very similar wood structure. While it does appear to be closely related to this genus, the drawings in Chun & Kuang (1958) show it to be sufficiently distinct for generic recognition.
The genus Tsuga as traditionally composed comprises a group of about nine very similar and closely related typical (type T. sieboldii Carr.) and two aberrant species, T. longibracteata Cheng and T. mertensiana (Bong.) Carr.; these two are each given generic status by Page (1988). Of these the former is much the more distinct, showing many clear similarities to Keteleeria and some to Pseudolarix, and is here accepted as a distinct genus with one species, Nothotsuga longibracteata (Cheng) Hu ex Page. The other species, T. mertensiana, is retained here in Tsuga, though in a distinct monotypic subgenus, subgen. Hesperopeuce (Engelm.) Ueno.
|
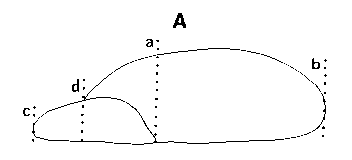
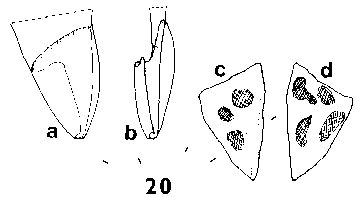
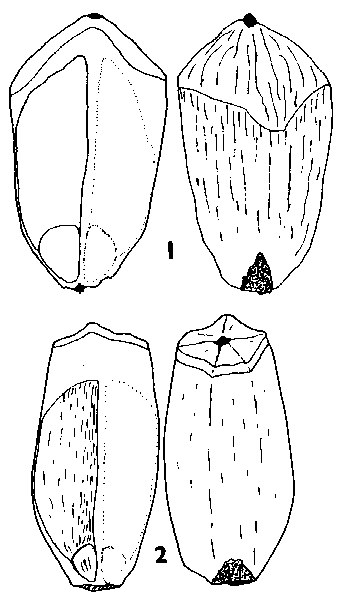
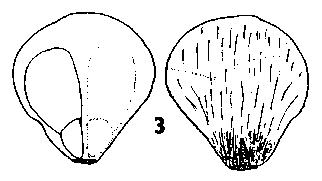
The seeds of a selection of species of the eleven genera are shown in Fig. 1 Four basic seed forms can be distinguished in the family, and are keyed out below, followed by a detailed description of the seeds of each genus. Fig. A. diagram indicating measurements used in this paper. Wing length given is from the seed tip, a-b, not d-b; wing to seed length ratio is a-b : a-c. |
|
1. Seed held in the wing by two 'claws', not in a cup; resin vesicles absent: Pinus only.
2. Seed field in a shallow cup on the wing, easily separated from the wing; fertile seeds dark, without resin vesicles: Picea only.
3. Seed field in a shallow cup on the wing, not separable from the wing; fertile seeds pale, without resin vesicles: Larix, Cathaya, Pseudotsuga.
4. Seed held in a deep cup on the wing, separable from the wing with difficulty; seeds with resin vesicles: Cedrus, Abies, Pseudolarix, Keteleeria, Nothotsuga, Tsuga.
|
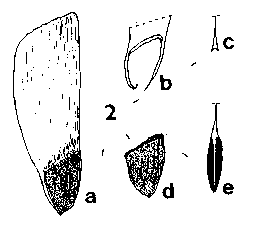
SEED FORM 1 Pinus. Seed very variable, small to very large, 3–26 mm long, without resin vesicles. Fertile seed commonly dark grey-brown to black, but pale brown, golden or freckled in a few species; infertile (hollow) seeds paler than fertile seeds, often whitish. Seed held in a two-clawed wing, these two claws always present even when the remainder of the wing is vestigial or absent (e.g. P. cembra L., P. edulis Engelm.). Wing separable or not separable from the seed, extending 0–2 mm (i.e., the thickness of the claws, in e.g. P. albicaulis Engelm.) to 35 mm (P. roxburghii Sarg.) beyond the end of the seed. In species with effective wings, wing colour always at least slightly, often strongly, streaked dark on pale brown; very flexible and easily bent without breaking; the base of the wing by the claws slightly to greatly (e.g. P. coulteri D. Don) thickened. Fig. 1.
Seed morphology in Pinaceae: abaxial and side views of selected species.
|

|
|
|
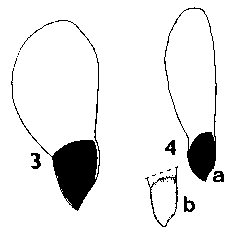
SEED FORM 2 Picea. Seed small, 3-6 mm, without resin vesicles; fertile seeds dark brown to black, infertile seeds paler, usually mid-brown. Seed held in a shallow cup, covered fully on the one side but not at all on the other; very easily separated from the cup. Wing always aerodynamically effective, extending 6–15 mm beyond the seed, pale, unstreaked, yellow- or pinkish-brown, moderately flexible, base not or only very slightly thickened. Ratio of wing : seed length fairly constant, about 2–3 in all species. Fig. 1. 3, Picea smithiana. 4, Picea farreri: a; b, base of wing showing cup. |
|
|
SEED FORM 3 Larix. Seeds small, 4–6 mm, without resin vesicles; fertile seeds whitish, lightly freckled pale brown; infertile seeds not differing. Seed held in a shallow cup, fully covered on one side and extending over the tip on the other side, firmly adnate to the cup and not separable. Wing always effective, extending 6–9 mm beyond the seed, commonly oriented at 20°–40° to the long axis of the seed; mid to dark purplish brown, often lightly streaked darker, and darker on the cup, moderately flexible; base slightly thickened and often bent. Fig. 1. 9, Larix potaninii: a; b, side view-note bend in wing. |
|
|
Cathaya. (After Chun & Kuang, 1958; Cheng & Fu, 1978). Seeds moderately small, 5–6 mm, pale, freckled darker, appearing similar to those of Pseudotsuga. Wing brown, streaked, darker on the cup, extending 10-15 mm beyond the seed. Fig. 1. 8, Cathaya argyrophylla (after Chun & Kuang, 1958). |

|
|
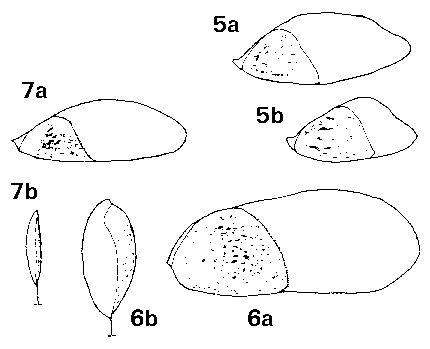
Pseudotsuga. Seeds medium, 7–9 mm or (in Ps. macrocarpa (Torrey) Mayr only) large, 12 mm, without resin vesicles; fertile seeds whitish, lightly freckled pale or mid-brown; infertile seeds not differing. Seed held in a shallow cup, fully covered on one side and extending over the tip on the other side, firmly adnate to the cup and not separable. Wing moderately to fully effective, extending 4–14 mm beyond the seed; length often variable with the position in the cone, markedly so in Ps. japonica (Shirasawa) Beissner: 4 mm wings in the small scales at the apex of the cone, 9 mm wings in the larger scales of the same cone. Wings pale to mid brown, often streaked darker brown, the cup darker, often red-brown; moderately flexible, straight (not angled nor bent at the base); base slightly thickened. Fig. 1. 5, Pseudotsuga japonica: a & b, two seeds from one cone. 6, Pseudotsuga macrocarpa: a; b, side view. 7, Pseudotsuga menziesii: a; b, side view. |
|
|
SEED FORM 4 Cedrus. Seeds large, 11–16 mm, grey-brown, fertile and infertile seeds not differing. Two to five resin vesicles on each side of the seed. Seed held in a deep cup, fully covered on one side, on the other side a flap covering about Ľ of the seed; the whole cup very thin and flimsy, fibrous, tearing easily off the seed. Wing fully effective, broadest near the tip, triangular, extending 15–25 mm beyond the seed; ratio of wing : seed length about 1.5 (higher than in Abies). Wing similar in colour to seed, unstreaked, moderately flexible, base slightly thickened. Fig. 1. 10, Cedrus libani. |
|
|
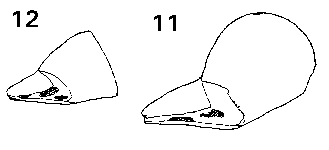
Abies. Seeds medium to large, 6-15 mm, mid- or grey-brown, or variously pigmented purple or violet, particularly before maturity, fertile and infertile seeds not differing. Two to five resin vesicles on each side of the seed. Seed held in a deep cup, fully covered on one side, on the other side a wide flap covering about Ľ–˝ of the seed; cup not thin or flimsy. Wing fully effective, broadest near the tip, triangular (except in A. bracteata, where rounded), extending 6–17 mm beyond the seed; ratio of wing : seed length about 0.8–1.2, fairly constant. Wing similar in colour to seed, unstreaked, moderately flexible, base slightly thickened. Fig. 1. 11, Abies bracteata. 12, Abies veitchii.
|

|
|
20, Abies pinsapo: a & b, two views of seedwing cup; c, abaxial view of seed; d, adaxial view of seed. |
|
|
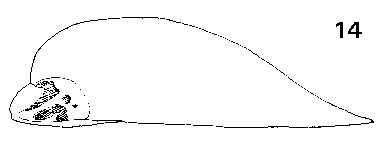
Pseudolarix. Seeds medium, 8 mm, buff to mid-brown, fertile and infertile seeds not differing. One to three resin vesicles on each side of the seed. Seed held in a deep cup, fully covered on one side, on the other side a flap covering about Ľ of the seed. Wing fully effective, broadest near the base, narrow pointed triangular, extending 18–27 mm beyond the seed; ratio of wing : seed length about 3, high to compensate for the narrowness of the wing. Wing similar in colour to seed, fairly thick, inflexible, very brittle, base slightly further thickened. Fig. 1. 14, Pseudolarix amabilis. |
|
|
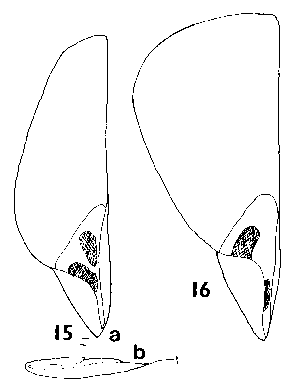
Keteleeria. Seeds large, 8–16 mm, buff to mid-brown, fertile and infertile seeds not differing. One to three resin vesicles on each side of the seed. Seed held in a deep cup, fully covered on one side, on the other side a flap covering about Ľ of the seed. Wing fully effective; in K. davidiana (Bert.) Beissner and allies, narrow, broadest below the middle (similar to Pseudolarix); in K. fortunei (Andr. Murray) Carrière and allies, much broader, broadest above the middle (intermediate between K. davidiana and Abies in shape). Wings in K. davidiana extending 14–22 mm beyond the seed with a wing : seed length ratio of 1.5–2; in K. fortunei about 13–19 mm and ratio about 1–1.2. Wing similar in colour to seed, fairly thick, inflexible, very brittle, base slightly further thickened. Fig. 1. 15, Keteleeria davidiana: a; b, side view. 16, Keteleeria fortunei. |
|
|
Nothotsuga. Seed moderately small, broad, 5–6x4 mm, mid-brown, fertile and infertile seeds not differing. Four to ten small resin vesicles on each side of the seed. Seed held in a deep cup, fully covered on one side, on the other side a flap covering about Ľ of the seed. Wing fully effective, broadest near the middle, round-ended, extending 7 mm beyond the seed; ratio of wing : seed length about 1.3. Wing similar in colour to seed, thin, inflexible, very brittle, base not thickened. Fig. 1. 17, Nothotsuga longibracteata. |
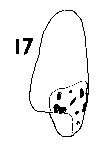
|
|
Tsuga. Seeds small, slender, 3–5 x 2–3 mm, pale brown or, in T. mertensiana only, often tinged pinkish; fertile and infertile seeds not differing. Numerous (5–20) very small resin vesicles on each side of the seed. Seed held in a deep cup, fully covered on one side, on the other side a flap covering about 1/8 – Ľ of the seed. Wing fully effective, broadest near the middle, round-ended, extending 5–8 mm beyond the seed; ratio of wing : seed length about 1.5–2. Wing paler than seed, buff, or often pink tinged in T. mertensiana, in all species often darker red-brown on the cup; very thin, moderately flexible but also brittle, base not thickened. Fig. 1. 18, Tsuga mertensiana. 19, Tsuga sieboldii: a; b, side view. |
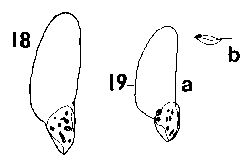
|
Mature cone scale morphology in Pinaceae is varied but divides into two major groups (Fig. 2), each further divided into smaller subgroups, separable by differences in the shape and growth of the ovuliferous scale (hereafter referred to as the scale) and the position of the seeds on it, and the size and form of the bract scale (hereafter, the bract). These sub-groups correlate well with the different groupings indicated by the seed morphology and are keyed out below, with brief descriptions of the cones, scales and bracts, concentrating on significant or poorly described features.
1. Scales with a broad, thick base, wholly covering the seed from abaxial view; the seeds lying in two depressions at the base of the scale.
1a. Cones biennial, rarely triennial; scales with a distinct umbo; bract small: Pinus only.
1b. Cones annual, scales without an umbo; bract small, rarely medium; cone peduncle very short, without leaves or bracts: Picea only.
1c. Cones annual, scales without an umbo; bract medium to long; cone peduncle moderately long, with leaves or leaf-like bracts: Larix, Cathaya, Pseudotsuga.
2. Scales with a slender, weak base, with the seeds extending well over the edge of the scale base and thus visible in abaxial view of the scale.
2a. Cones biennial; scales without a distinct umbo; bract small; cone peduncle very short, without leaves or bracts: Cedrus only.
2b. Cones annual; scales without an umbo; bract medium to long; cone peduncle short to long, with or without leaves or leaf-like bracts: Abies, Pseudolarix, Keteleeria, Nothotsuga, Tsuga.
|
CONE FORM 1a Pinus. Cones spreading or pendulous, failing at or soon after maturity, or long persistent, short to long pedunculate with minute scale leaves on the peduncle; biennial or (in 2-3 species) triennial. Scales broad based, always bearing an umbo, the result of interrupted growth between the growing seasons. Very variable in consistency, mostly thick and woody, in some species thinly woody and in others thick but soft and not woody. Bracts short, thick and irregular in shape, rough and matt. Fig. 2. Cone scale morphology in Pinaceae.
|
|
|
CONE FORM 1b Picea. Cones pendulous, failing soon after maturity or persistent 2-4 years, sessile, without leaves or bracts at the base. Scales broad based, thinly woody, tough, strongly attached to the cone rachis, always at least a little longer than wide. Bracts short, thin, simple acute or somewhat irregular, mostly rough and matt, occasionally smooth and lustrous. Fig. 2. 3, Picea breweriana. |
|
|
CONE FORM 1c Larix. Cones erect, annual, long persistent after maturity; pedunculate with numerous leaves grading into bracts in the peduncle. Scales broad based, very tough, coriaceous and strongly attached to the rachis, usually a little wider than long. Bracts medium to long, trilobate or aristate-acuminate, thin, smooth, lustrous or bloomed. Fig. 2. 6, Larix griffithiana. |
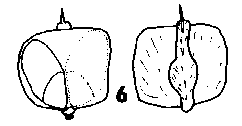
|
|
Cathaya. Cones spreading, annual, long persistent after maturity, short pedunculate with modified leaf-like bracts on the peduncle. Scales broad based, tough, a little wider than long. Bracts moderately long, simple acuminate, smooth. (Chun & Kuang, 1958.) Fig. 2. 5, Cathaya argyrophylla (after Chun & Kuang, 1958). |
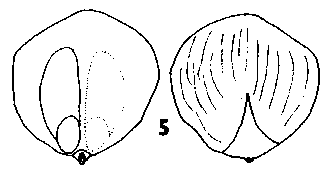
|
|
Pseudotsuga. Cones pendulous, annual; in the Asiatic species long persistent; in the American species falling soon after maturity, short pedunculate with modified leaf-like bracts on the peduncle. Scales broad based, very tough, coriaceous and strongly attached to the rachis, always at least a little wider than long. Bracts long to very long, tridentine, thin, smooth and lustrous. Fig. 2. 4, Pseudotsuga menziesii. |
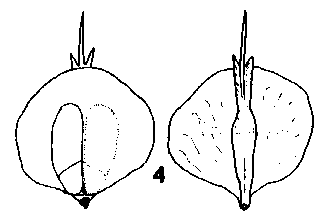
|
|
CONE FORM 2a Cedrus. Cones erect, biennial, disintegrating at maturity by abscision of the scales from the rachis, sessile without leaves or bracts at the base. Scales hard, tough and woody before maturity, becoming loose and easily broken after, narrow based broad peltate, covered with dense long pubescence except at the base and exposed apex. Bracts very short, acute, not showing further growth during the second growing season while the scales are still growing. Rachis persistent on the branch. Fig. 2. 7, Cedrus atlantica. |
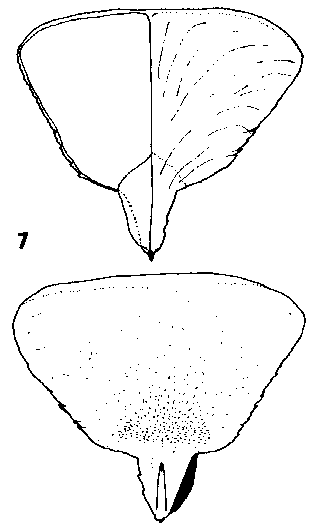
|
|
CONE FORM 2b Abies. Cones erect, annual, disintegrating at maturity by abscision of the scales from the rachis, sessile or with a short leafless peduncle. Scales soft, easily broken, not tough, narrow based broad peltate, often auriculate, variably short pubescent. Bracts medium to very long, in A. bracteata extremely long; commonly trilobate with two broad lobes either side of a slender central point, but in some species (e.g. A. magnifica Andr. Murray) the lobes reduced and approaching simple acute; thin, smooth, lustrous or bloomed. Rachis persistent on the branch. Fig. 2. 8, Abies grandis. |
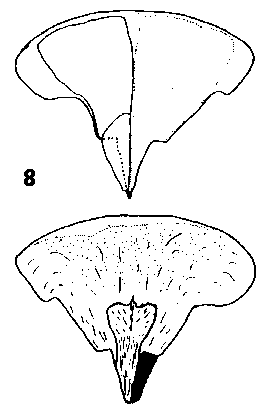
|
|
Fig. 2. 9, Abies bracteata. |
|
|
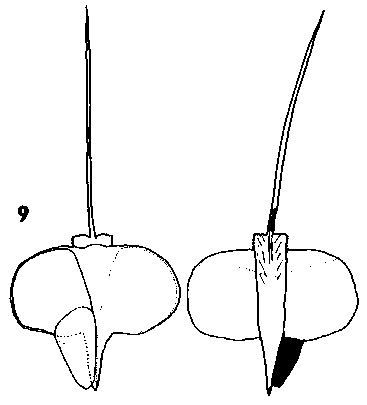
Pseudolarix. Cones erect, annual, disintegrating at maturity by the breakup of the rachis, moderately long pedunculate with a leafy peduncle. Scales soft, easily broken up, not tough, narrow based, narrow acute triangular, auriculate, with scattered short pubescence (under lens). Scale apex spreading, with the seedwings exposed before maturity. Bracts medium, acute or slightly lobed, thin, smooth, lustrous. Rachis not persistent, breaking up with the cone. Fig. 2. 10, Pseudolarix amabilis. |
|
|
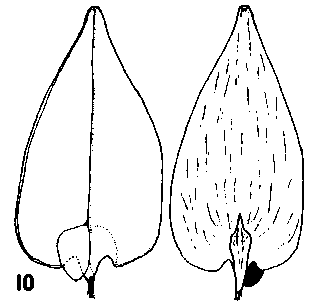
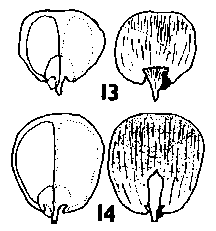
Keteleeria. Cones erect, annual, usually breaking up on the tree within a year of maturity by the disintegration of the rachis, or falling complete in a variable state of disrepair; long pedunculate with a leafy peduncle. Scales soft, easily broken up, not tough, narrow based, auriculate, pubescent. Shape variable; in K. davidiana and allies, much as in Pseudolarix though broader and with a rounded apex, the apex spreading as in Pseudolarix: in K. fortunei and allies, more broad peltate, similar to Abies though with a more rounded (less obtriangular) apex, the apex not spreading before maturity. Bracts medium, trilobate with small lobes, thin. smooth, lustrous. Fig. 2. 11, Keteleeria davidiana. |
|
|
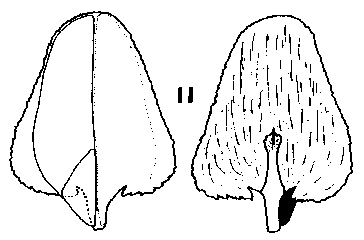
Nothotsuga. Cones erect, annual, persistence not recorded, long pedunculate with a leafy peduncle. Scales soft, not tough, narrow based, auriculate, moderately broad peltate, apex not spreading before maturity. Bracts long, acute or weakly trilobate, thin, smooth, lustrous. Fig. 2. 12, Nothotsuga longibracteata. |
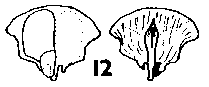
|
|
Tsuga. Cones pendulous, annual, falling complete shortly after maturity, a little later in T. mertensiana, sessile, leafless. Scales soft, not very tough, narrow based but with a very short base, auriculate, narrow peltate to round, glabrous or thinly pubescent, except in T. mertensiana which has dense, velvety short pubescence. Scale apices not spreading. Bracts short, rounded, in T. mertensiana trilobate and a little larger, thin, smooth and lustrous. Fig. 2. 13, Tsuga sieboldii. 14, Tsuga mertensiana. |
|
The male cones of the Pinaceae are not easy to study, being available for only a very limited period in the year, often inaccessible as fresh material and not easily nor commonly preserved as herbarium material (Page, 1979); it has therefore not been possible to study these very extensively for this paper. They do however exhibit some features of value in determining relationships. In most of the genera they occur individually, with one in each bud and separate from any foliage. In Pinus they are clustered 10–150 cylindrically round the young stems arising from one shoot bud with the foliage, though also individually separate in small buds on this shoot as well.
In three genera, Pseudolarix, Keteleeria and Nothotsuga, the male cones occur in umbels of 5–20 arising together from a single bud (Cheng & Fu, 1978), unusual in Pinaceae though shared by some members of Taxodiaceae (Sciadopitys and Cunninghamia) among the other conifers.
In Abies the male cones are grouped 5–50 in close proximity on the undersides of a relatively limited number of shoots (see Phillips, 1978:64 for photographs), while in the remaining genera they are much more scattered, rarely occurring in pseudowhorls of more than five or six male cone buds on a single shoot. This last also applies to the buds of the umbellate genera.
The morphology of the individual male cones is broadly similar throughout the family, varying mainly in size. Those of Cedrus are the largest, 5 cm expanding to 8 cm after shedding pollen; Larix and Tsuga the smallest, under 1 cm when expanded; the remainder mostly 3–5 cm. Larix differs from the other genera in having relatively broader cones, globular rather than ovoid before pollen release, but the difference is minor. Those of Cedrus shed pollen in autumn, the other genera in spring.
The pollen is either unwinged or winged with two air bladders; Liu (1971:58, table 3) and Page (1988) detail the occurrence of these two forms.
The female cones at pollination stage may also provide important evidence of relationships, but it was not possible to obtain adequate material for any useful study. Sporne (1974:139) records that the female cones at pollination of Pinus and Picea contain micropylar fluid, absent in the other genera.
The foliage of the Pinaceae genera is well described and has previously been considered to be of great importance in the determination of generic relationships (e.g. Melchior & Werdermann, 1954). However, the importance of the tree's foliage to its growth and success in competition, and the necessary exposure of the foliage for maximum photosynthetic advantage render it particularly liable to evolutionary pressure. Its features should therefore be viewed with caution generally, and in this the Pinaceae is no exception. In particular, the separation of subfamilies on the basis of the occurrence of short shoots is probably artificial. It is clear from the widespread occurrence of such shoot dimorphism not only in Pinaceae but also in other widely separated groups of Gymnosperms (e.g. Sciadopitys, Ginkgo, Phyllocladus) and Angiosperms (e.g. Berberis and Cotoneaster to name two of many) that under certain conditions it can give a plant great evolutionary advantage, and also that it has arisen on many occasions and may thus only represent convergent evolution within the Pinaceae.
It should be noted that all of the Pinaceae genera show some dimorphism in their shoots to a greater or lesser degree. The stylized representation of two-year twigs of some genera in Fig. 3 shows that although the dimorphism grades from strong to weak in different genera, none show true monomorphism with all shoots of equal development.

Fig. 3. Dimorphism variation in Pinaceae.
Greatest in Larix, least in Abies; theoretical monomorphic shoot, right, with all growth of equal length.
FOLIAGE STOMATA
The distribution of stomata on the leaves appears to be correlated primarily with climate; species from high rainfall areas tend to have no stomata on surfaces facing upwards and dense, very wax-rich bands of stomata below (e.g. Abies grandis (Douglas) Lindley, Picea farreri Page & Rushforth, Pinus monticola Douglas), while species from drier climates tend to have stomata on both surfaces (e.g. Abies concolor (Gord.) Lindley, Picea asperata Mast., Pinus sylvestris L.), though many exceptions occur. This stomatal distribution appears to be highly adaptive, with closely related taxa often separated into these two categories (e.g. Abies grandis/A. concolor). In most species where stomata occur on both surfaces, one surface has a denser and more regular distribution of stomata, this corresponding with the surface on which they are found in those species of the same genus where they only occur on the one side. Exceptions are a number of Pinus and Picea species in which the distribution is equal on both surfaces, e.g. Pinus nigra Arnold.
These more obvious stomatal bands, here termed primary stomatal bands, are found on the abaxial surface of the leaf in most Pinaceae genera, but are adaxial in those species of Pinus and Picea where they are not equally distributed. No selective pressure can be found to account for this difference in strategy.
The leaves of Pinus and Picea also differ from the other genera in not having a distinct petiole, a factor also unlikely to have derived from selective pressure.
FOLIAGE AROMA
The fresh foliage of most species in the family is aromatic, either continuously or on being crushed; in Pinus, Picea, Abies, Cedrus and some Pseudotsuga species, it is fruity- or sour-resinous, with a 'Christmas' smell; in Larix and the other Pseudotsuga species, there is a slight hay (cournarin) scent from the crushed leaves but a more resinous scent in the bark of the twigs. Tsuga, Keteleeria and Pseudolarix have no resinous scent, either in the foliage or the wood; different Tsuga species smelling of various umbelliferous species (Mitchell, 1974; also T. mertensiana parsley-scented; pers. obs.) and the other two genera having a cucumber- or avocado pear-like scent. The foliage aroma is persistent on drying in only a few strongly aromatic species (e.g. Abies grandis) and is lost in herbarium material; the scents of Cathaya and Nothotsuga remain unknown. A strong and very pleasant aroma is retained for many years in the heartwood of Cedrus and also in the cones of Tsuga mertensiana and a few pines, notably Pinus cembroides Zucc.
Pinus & Picea
The placing of Pinus in a monogeneric subfamily by Pilger (1926) on the basis of its unique cone, seed, shoot and leaf morphology is fully justified. Its closest relative appears to be the morphologically very distinct Picea, with which it shares a number of features not found in the other genera. The most important of these is probably the presence of micropylar fluid in the female cone, which probably indicates a divergence from the other genera at a very early stage. The position of the primary stomatal bands, absence of an obvious leaf petiole, the often similar form of the bract and the dark seeds with a colour difference between fertile and infertile seeds gives further evidence for such a relationship. Picea does also show similarities to other genera, notably the broad base to the cone scale, shared with Larix, Pseudotsuga and Cathaya as well as with Pinus, but it differs from these in seed morphology as well as in the features given above. It thus appears, like Pinus, to have no very close relatives.
Larix, Cathaya & Pseudotsuga
These three genera are very similar to each other in seed and cone scale structure. The broad base of the cone scale is shared with Pinus and Picea, but the bract is much more similar to some of the remaining genera, notably Abies, Keteleeria and Pseudolarix, to which their foliage is also closely similar. Their seed is however very distinct from these and all other genera in the strongly adnate wing and pale freckled testa. The position of Cathaya in this group must remain somewhat open, but the excellent detail shown in Chun & Kuang (1958) strongly indicates its inclusion here. In many respects it appears to be intermediate between Larix and Pseudotsuga, probably closer to the latter. It has noticeably dimorphic shoots, though less so than Larix and the cones are on the longer, not the shorter, shoots. The cone is closely similar to the asiatic species of Pseudotsuga in all aspects except for the shorter, unlobed bracts. It has also been considered close to Pseudotsuga and Larix in its wood structure by Xie (1957; see Page, 1988) and by Greguss (1970) who actually incorporated Cathaya in Pseudotsuga, though publishing the combination invalidly. Greguss also pointed out that the similarity to Picea found by Yatsenko-Khmelevsky & Budkevich (1958) is erroneous.
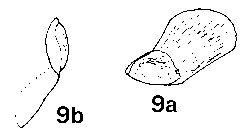
The evidence here does not support the recent incorporation of Cathaya in Tsuga by de Laubenfels & Silba, given on the basis of chromosome number, foliage arrangement and similar pulvini (Silba, 1984). Its chromosome number of 2n = 24 is not different from the vast majority of the Pinaceae–only two species are known to differ, Pseudotsuga menziesii (Mirb.) Franco (2n = 26) and Pseudolarix amabilis (Nels.) Relid. (2n = 22) (Rushforth, 1987:196); while plates 9 & 10 in Chun & Kuang (1958) show a minimal degree of resemblance to Tsuga. The pulvini are equally similar to Larix and the foliage arrangement is very similar to that shown by young plants of Pseudotsuga wilsoniana Hayata grown by the author.
The apparent close similarity of the foliage of Larix to Pseudolarix does not extend to any great detail and is confined to the shoot arrangement and deciduous leaves–both factors very liable to selective pressure–and is almost certainly due to convergent evolution. The shoot dimorphism of Pseudolarix is noticeably less than that of Larix, with the short shoots extending about 3 mm annually as opposed to under 0.5 mm in Larix. The pulvini of Larix also differ, being much more pronounced and decurrent along the shoot. There is no close similarity in the cone or seed morphology, with Pseudolarix showing much closer relationships elsewhere.
A possible relationship between Pseudotsuga and Tsuga is discussed later in the paper.
Cedrus, Abies, Pseudolarix, Keteleeria, Nothotsuga & Tsuga
These remaining six genera all share a highly distinctive seed and cone scale morphology and are clearly related. Only Cedrus shows any important deviation in detail, with its thick woody scales and very small bracts, probably consequential from the longer period of cone maturation, c. 24 months versus 6–8 months. It still shares the narrow scale base and distinctive seed morphology.
The seeds of all of these six genera have resin vesicles on the testa. This resin has a rather unpleasant smell and taste (pers. obs.!), similar in all six genera, and is probably a defence against predation by birds and mammals. Cones of' these genera (Tsuga less so) are only rarely attacked by squirrels (Sciuridae) and crossbills (Loxia), the chief vertebrate predators on the seeds of the other genera, though certain nuthatches (Sitta) do appear to take the seeds regularly (van der Berg, 1985). This chemical defence may be connected with the softness of the cones and, in five of the six genera (not Tsuga), the tendancy of the cones to be carried on stouter branches (more easily able to carry a squirrel) than in Picea, Larix, Pseudotsuga and (?) Cathaya (but not Pinus). The resin vesicles are scattered randomly over the seed, the pattern varying even from seed to seed in one cone, though the number of vesicles appears to be more or less regular within each genus.
The seeds are also distinct in the depth and coverage of the cup, with a flap extending over part of the abaxial side; this flap coincides exactly with the extension of the seed over the edge of the scale stem (see Figs 1 & 2) and is probably to give extra protection to the seed during development. In Cedrus, where the thick cone scales are arranged very tightly, giving better protection, this flap and the rest of the cup is very thin and weak, usually disintegrating if removed from the seed.
Cedrus & Abies. Within this group of six genera, Cedrus and Abies appear to be most closely related to each other, despite the sometimes anomalous morphology of Cedrus. They are unique in the well-developed cone scale abscision and in leaving the cone rachis persistent on the branch; they also have similar broad peltate cone scales, and differ from the remaining four genera in the resinous aroma of the foliage, and (except Tsuga) in the short or very short and leafless cone peduncle.
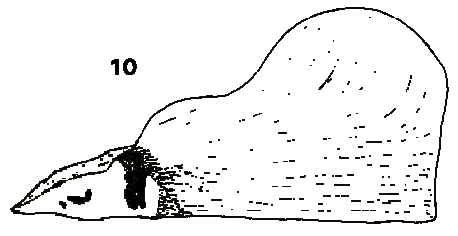
The species Abies bracteata is very interesting and distinct, and the possibility of separate generic recognition should be considered. The cone is unique in the whole Pinaceae in the extraordinary elongation of the bracts, exserted up to 4 cm beyond the scales, and is distinct from other Abies in the shape of the scale and seedwing (see Figs 1 & 2). The bud is also unique, long, slender and sharp pointed with parchment like scales, somewhat resembling the buds of Pseudotsuga though larger and different in texture and colour (buff, not red-brown), while the foliage bears some resemblance to (particularly juvenile) Keteleeria foliage in the broad flat needles with a long acuminate spine tip. This resemblance is however probably due to convergent evolution in similar hot dry climates; other Abies species from a similar climate type (notably A. firma Sieb. & Zucc.) show similar adaptations, though to a lesser degree. While the resemblances to other genera are few and of doubtful significance, those to the remainder of Abies are great. The maturation and disintegration of the cone are identical; the cone rachis closely resembles that of A. fargesii Franchet; the cone peduncle, while the longest of any Abies, is closely approached by that of A. delavayi Franchet; the rhombic shape of the exposed portion of the cone scale also resembles that of these two species and the ovoid cone shape (disregarding the bracts) is shared by A. fargesii and A. firma. The preference of the author is therefore to recognize its distinction only at subgeneric level within Abies, as subgenus Pseudotorreya (Hickel) Franco, as also treated by Liu (1971) in his monograph of Abies.
Pseudolarix, Keteleeria & Nothotsuga. These three genera share a number of unusual and interesting features and appear to be closely related. They have umbellate male cones, unique in Pinaceae, and have cones set on long leafy peduncles. The cones are also semi-dehiscent, with the scales falling by disintegration of the rachis, not by abscision of the scales as in Cedrus and Abies. This occurs at seed maturity in Pseudolarix, but later in Keteleeria and Nothotsuga, the cones of which gradually break up some time after seed dispersal, usually falling to the ground in a half-disintegrated state.
There are two distinct cone forms in Keteleeria which serve well to show the affinities of the genus. The first, exemplified by the type species, K. fortunei, has adpressed scales which are widest above the middle, resembling many Abies and, more closely, Nothotsuga; while the second, exemplified by K. davidiana, has spreading scales, widest below the middle and free at the tips, exposing part of the seedwing, resembling Pseudolarix closely. The bracts are similar in both groups.
The unique hypogeal germination and coppicing ability of Keteleeria (Rushforth, 1987) may be best considered as a relatively recent adaptation to fire or grazing pressures in its dry climate and of limited value in determining relationships, though it would be interesting to know if this is shared by Nothotsuga, which has not been recorded.
The new genus Nothotsuga Hu ex Page (Page, 1988) is based on a species previously classified in Tsuga, T. longibracteata Cheng (Cheng, 1932). Herbarium specimens studied, Y. Tsiang 7712 (E. isotype) and Rushforth 404 (E), and the illustration in Cheng & Fu (1978) however indicate a very close relationship to Keteleeria as outlined above in the umbellate male cones and erect pedunculate cones with peltate scales broader than typical of Tsuga. The bract more closely resembles that of Pseudolarix than either Keteleeria or Tsuga in shape, but is relatively longer than any of these. The general aspect of the whole tree is most similar to Keteleeria (Rushforth, 1987).
Nothotsuga does also at the same time share many features with Tsuga; the shoot and pulvinus form, twig layout and pendulous leading shoots are close to those of Tsuga, the leaves have a sunken midrib as in Tsuga (ridged in Keteleeria and Pseudolarix). The cone size, seedwing shape, seed size and resin vesicle distribution are all intermediate between Keteleeria and Tsuga, but closer to Tsuga.
The species thus has very close affinities to both of these genera, yet does not fit well in either, and is therefore best treated as a separate genus. It has been considered by Van Campo-Duplan & Gaussen (1948) to be a recent natural hybrid between the two genera; this is not only highly improbable, but also does not account for either the Pseudolarix-like bract or the unequal level of resemblance between the different features. A far more likely explanation is that it is a relict from the period of the evolutionary divergence of this group of genera. The existence of fossils resembling this species (Page, 1988) would support this.
Tsuga. This is the most distinctive genus in this group. It obviously belongs here, with resin vesicles on the seed, an allaxial wing-cup flap and narrow based atiriculate cone scales and clear affinities to Nothotsuga. Yet it also shows some relationship to Pseudotsuga; the resin vesicles are very small and numerous, and contain little resin. Smaller, more numerous still and without resin, they would much resemble the freckling on Pseudotsuga seeds. The abaxial wing-cup flap is small, only a little larger than the small amount of wing cup extending on the abaxial side of Pseudotsuga seed, and in a similar position (see Fig. 1). Further, the seed wing is less easily separable from the seed than in the other five genera above, and is shaped much like that of Pseudotsuga. The cone scale has a shorter narrow stem than the other five, giving a more nearly broad based appearance; the cone is also the toughest and most persistent of the above group, and is pendent on slender shoots as in Pseudotsuga. The leaves are softer than the other evergreen members of its group, again resembling Pseudotsuga. It would thus appear that Tsuga represenis a form which has existed with relatively little morphological change ill these features since tile evolution of this group away from a probable cominon ancestor with Pseudotsuga and its allies.
One species of Tsuga, T. mertensiana, differs from other Tsuga species in many respects. It has in particular an atypical long cylindric cone, dense short pubescence on the cone scales, and thick needles, with stomata on both faces, carried on shoots which are more conspicuously sub-dimorphic than those of other Tsuga. The foliage shows much apparent similarity to Cedrus atlantica (Endl.) Manetti ex Carr., though is much softer. Page (1988), in pointing out these and other differences-notably the pollen structure-from Tsuga s.str., and observing the similarities to Cedrus and other genera in some features, proposed the segregation of this species under the old generic name Hesperopeuce Lemmon, as H. mertensiana (Bong.) Rydberg. It has also been proposed that T. mertensiana is a product of recent hybridization between T. heterophylla (Raf.) Sarg. and Picea sitchensis (Bong.) Carr. (Van Campo-Duplan & Gaussen, 1948).
All examination of the scales and seeds of this species reveals a very close relationship to Tsuga s.str.; the seeds and seedwings are indistinguishable except for a variable violet cast in the colour, and the individual scales in shape are within the range of variation of the rest of the genus, differing only in the velvety pubescence noted above. No other two genera in the family show this level of similarity. There is also no indication of any influence of Picea morphology; the hybrid origin theory is therefore rejected here.
The similarities to Cedrus do not extend to the cone morphology beyond what is shared by the other four genera in this group, and are confined to features noted previously as particularly liable to evolutionary pressure and thus could well be due to convergent evolution. Both species occur in regions of very high winter snowfall and high summer light intensity at high altitude. T. mertensiana does show more than the rest of the genus the above noted similarities to Pseudotsuga; it has in addition the longer cylindrical cone and the slightly trilobed as opposed to simple rounded bracts, and as the only Tsuga with adaxial stomata on the leaves, a particular resemblance to Ps. menziesii var. glauca (Beissn.) Franco, though this is also the only Pseudotsuga showing this feature, which may be due to convergent evolution.
In view of the considerably closer relationship of T. mertensiana to Tsuga s.str. than to any other Pinaceae genus, but also recognizing the differences given above and in Page (1988), the most suitable treatment for this species would be to retain it in Tsuga, but as a well-defined and distinct monotypic subgenus, Tsuga subgen. Hesperopeuce (Engelmann) Ueno (Ueno, 1957).
A further important consideration for retaining T. mertensiana in Tsuga is the existence of a taxon intermediate, and almost certainly hybrid between, T. mertensiana and T. heterophylla. This plant, T. xjeffreyi (Henry) Henry, occurs frequently in the wild where the ranges meet (Bean, 1980) and also in cultivation, though there appear to be no recorded instances of verified F1 hybrids (Taylor, 1972). All plants observed have shown very slow growth with no evidence of heterosis, and are probably the result of hybridization many generations previously. Seed collected from a specimen of T. mertensiana surrounded by T. heterophylla at Cragside, Northumberland was planted in October 1987, but the resulting seedlings are not yet large enough for hybrids to be identified (October 1988). It is worth noting that T. mertensiana is very much the dominant parent in the morphology of the hybrid, both in the cones and in the foliage (pers. obs.; Mitchell, 1974), possibly as a result of the slow growth limiting survival to areas at high altitude (where T. mertensiana is dominant) where there is less competition.
It has already been pointed out that the subfamilial divisions previously accepted are artificial. The evidence here suggests the existence of four sub-families in the Pinaceae; these are presented formally below with a synopsis of the genera contained. A possible evolutionary sequence for the family is given in Fig. 4.
|
Fig. 4. Diagram illustrating a possible phylogenetic sequence for Pinaceae as suggested by this study. The development of selected features is indicated 1-10 : 1, divergence of groups with and without micropylar fluid, ad-/abaxial primary stomatal bands and differing bract form; 2, two year cone maturation; 3, dwarf shoots and foliage dimorphism; 4, seed resin vesicles and narrow-based cone scales; 5, cone scale abscision; 6, leafy pedunculate cones; 7, erect mature cones; 8, deciduous foliage; 9, increasing shoot dimorphism; 10, more extreme shoot dimorphism. |

|
| 1. | Leaves strongly dimorphic, with adult foliage consisting of non-photosynthetic scale leaves and also photosynthetic leaves; the latter on normally sterile and ultimately deciduous dwarf shoots. Cones biennial, rarely triennial, with each year's scale growth distinct, forming ail umbo on each scale. Seedwing large or vestigial, holding the seed in a pair of claws | (Subfamily Pinoideae) Pinus | |||||||||||||||||||||||||||||||||||||||||||||
| + | Leaves not or only weakly dimorphic, always photosynthetic, on long or short, but not sterile dwarf shoots. Cones mostly annual, or if biennial, without a distinct umbo. Seedwing always large, holding the seed in a cup | 2 | |||||||||||||||||||||||||||||||||||||||||||||
| 2. | Seed without resin vesicles. Cone scale base broad, concealing the seeds fully from abaxial view | 3 | |||||||||||||||||||||||||||||||||||||||||||||
| + | Seed with resin vesicles. Cone scale base narrow, with the seeds partly visible in abaxial view | (Subfamily Abietoideae) 6 | |||||||||||||||||||||||||||||||||||||||||||||
| 3. | Mature fertile seed dark brown to black, easily separated from the seedwing. Leaves with primary stomatal bands adaxial (above the xylem) or equally on both surfaces | (Subfamily Piceoideae) Picea
| + |
Mature fertile seed pale brown to whitish, not separable from the seedwing. Leaves with primary stomatal bands abaxial (below the phloem vessels) only
| (Subfamily Laricoideae) 4
| 4. |
Leaves deciduous, shoots strongly dimorphic, cones erect .
| Larix
| + |
Leaves evergreen, shoots not or only weakly dimorphic, cones pendulous or spreading
| 5
| 5. |
Cone bracts moderately short, simple acuminate.
| Cathaya
| + |
Cone bracts long, tridentine
| Pseudotsuga
| 6. |
Mature cones pendulous
| Tsuga
| + |
Mature cones erect
| 7
| 7. |
Mate cones solitary (one per bud). Seedwing tips not exposed on the mature cone, cone scales neatly deciduous leaving the cone rachis persistent on the branch
| 8
| + |
Male cones umbellate. Seedwing tips often partly exposed on the mature cone, cone scales deciduous or subpersistent, the rachis breaking up or falling with the cone
| 9
| 8. |
Cones mature in 7 months, soft. Shoots not dimorphic
| Abies
| + |
Cones mature in 24 months, hard, woody. Shoots dimorphic
| Cedrus
| 9. |
Leaves deciduous. Shoots strongly dimorphic. Cone scales deciduous
| Pseudolarix
| + |
Leaves evergreen. Shoots weakly dimorphic. Cone scales subpersistent
| 10
| 10. |
Cones large, over 6 cm. Pulvini not prominent. Shoots not pendulous
| Keteleeria
| + |
Cones small, under 6 cm. Pulvini prominent. Leading shoots pendulous
| Nothotsuga
| |
This study would not have been possible without the generous help of many people. My thanks are due to those who provided study material, often at short notice and in difficult circumstances after the October 1987 storm in SE England, particularly David Capewell (Horsham), Jim Keesing (Kew) and Tony Schilling (Wakehurst Place); to Soren Odum (Horsholm Arboretum, Denmark) and the International Dendrology Society for allowing me to present an early draft of this paper at the IDS Spruce & Fir conference, August 1986; and most of all to Keith Rushforth (Portsmouth) and Chris Page (Edinburgh) for their considerable advice, encouragement, and assistance in carrying out this study.
The author would be most grateful to receive material, particularly cones and seeds, of Cathaya argyrophylla and Pinus krempfii Lecomte for further study.
He would also make a plea, to any author wishing to give generic distinction to Abies bracteata, not to use the name 'Pseudotorreya' for this purpose; two 'Pseuds' in the family is quite enough!
YATSENKO-KHMELEVSKY, A. A. & BUDKEVICH, E. V. (1958). On the wood anatomy of Cathaya argyrophylla.